原文标题为:Still waters run deep: Large scale genome rearrangements in the evolution of morphologically conservative Polyplacophora(静水流深:形态保守的多板纲在进化中的大规模基因组重排)

摘要
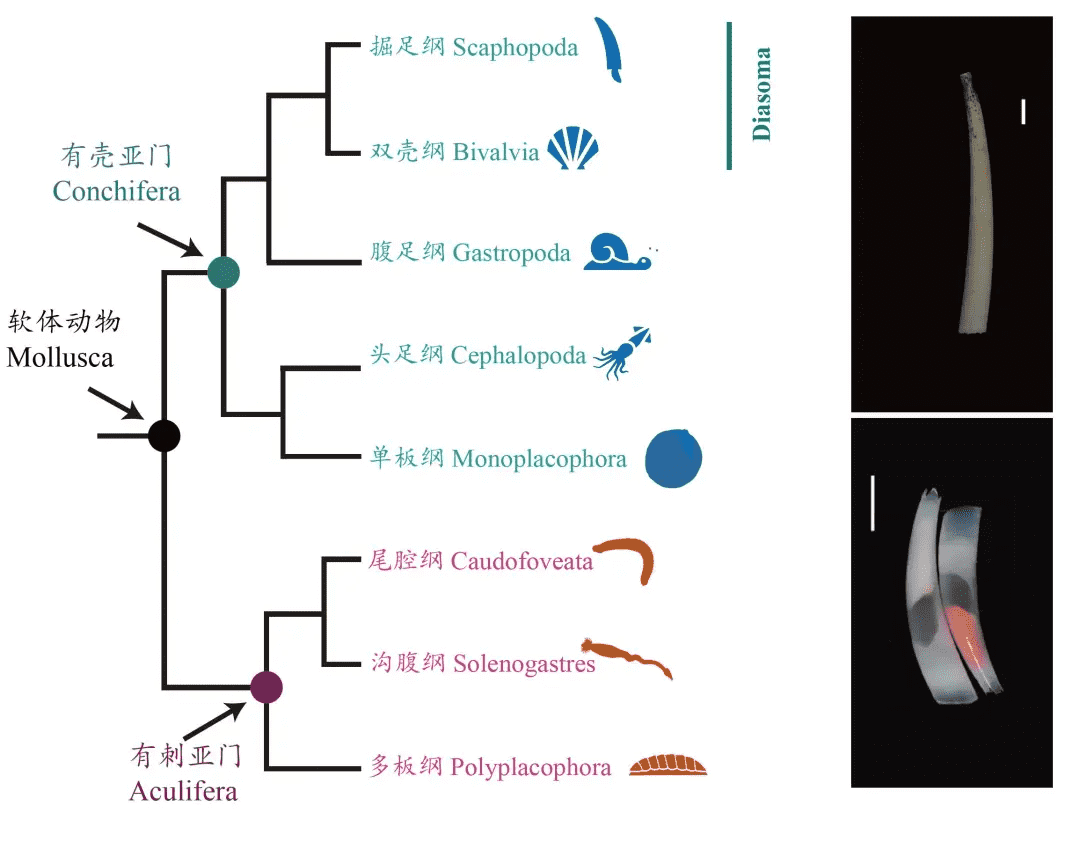
软体动物门(Mollusca)分为超高多样性的有壳亚门(Conchifera)(包括腹足纲、双壳纲、头足纲等)和有刺亚门(Aculifera)(多板纲和无板纲)。对于深度分化、形态上保守的石鳖类(chitons)而言,高质量的基因组数据提供了探索跨越动物进化长时间跨度的古老遗传特征和进化机制的机会。我们进行了五份参考质量基因组的比较分析,包括四份新组装的基因组,涵盖了所有主要的石鳖类群:侧鳞石鳖目(Deshayesiella sirenkoi)、Callochitonida目(Callochiton septemvalvis)和石鳖目(Acanthochitona discrepans和A. rubrolineata),共四个物种。并更新了软体动物的系统发育树。与有壳亚门基因组的比较分析,发现了20个古老的软体动物连锁群(MLGs),这些连锁群在双壳纲染色体中相对保守,但在石鳖类和有壳亚门基因组中经历了重排、重组、融合和早期的重复(或裂变)。两个同属的物种各有8条染色体(单倍体,2n=2x=16),分别来自其最近共同祖先染色体的两次独立融合事件。最大数量的新融合出现在被认为是最原始类群的侧鳞石鳖目(Lepidopleurida)中,而石鳖目日本花棘石鳖(Liolophura japonica)具有部分基因组重复,已知的软体动物内大规模基因重复的发生范围更加广阔。该纲极端而动态的基因组重排与大多数其他动物形成鲜明对比,并不能用该类群久远分化时间以来的累积变异来解释。石鳖类表型上的保守性状并未在其基因组中体现出保守性。
Song et al., 2023 PNAS
背景介绍
推动或限制重大创新演化的基因组机制仍然是进化生物学的要解决重要问题之一。现存的软体动物是各动物门中形态差异最大的动物类群:软体动物的身体构造(body plans)包括鱿鱼、蠕虫、活石和糖果色的树蜗牛(squid, worms, living rocks and candy-coloured tree snails),以及化石记录中已知的各种中间和新奇的形式。早期在减数分裂中对染色体数量的研究使用了软体动物,对跨物种染色体数量模式的理解也为动物多倍化提供了基本见解。尽管在早期进展中发挥了关键作用,但这个重要的动物门在全基因组序列数据的质量和分类覆盖范围方面仍然落后于其他动物门。重建多样化的软体动物的基因组结构演化历史,对于理解动物的基因组演化至关重要。
先前的研究合理地假设在主要类群内,染色体内基因易位的速率是恒定的。如果这是真的,共线性重排可以作为分歧时间的钟状指示器。但在软体动物中,基因组重排的速率尚不明确,也不清楚在这个庞大的类群中如重排速率何变化,而较高的重排速率会搅乱祖先状态重建。在软体动物中,良好的化石证据提供了分歧时间的独立记录,这将为重排速率作为分歧时间度量的可变性(和实用性)提供更多见解。
石鳖类长期以来被认为是理解软体动物祖先身体构造的关键类群。全球约有1000种现存的石鳖物种,它们都拥有一个八部分组成的壳甲,自石炭纪以来的3亿多年,其在表型上几乎没有变化,变异较小。石鳖在仿生设计中越来越重要,这将需要我们利用基因组工具,以了解其柔性甲胄、独特感官系统和铁矿化齿舌(iron mineralised radula)的遗传调控机制。石鳖雌雄异体,大多数通过广播产卵(broadcast spawning)繁殖,是食草者,大多数没有生态特化。石鳖通常是保守的,但已知的石鳖物种丰富度比形态和行为上更为多样的头足类更高。驱动这一类群物种形成的适应性机制仍是一个未解的问题。
在此,我们测序了覆盖现存石鳖三大类群的四个新参考质量的基因组:Deshayesiella sirenkoi(侧鳞石鳖目)、Callochiton septemvalvis(Callochitonida)、Acanthochitona discrepans和A. rubrolineata(石鳖目)。我们将这些基因组与先前发表的基因组数据结合使用,重建了软体动物门的系统发育树。这使我们能够自信地重建软体动物以及多板纲内部不同的祖先染色体重排事件。
结果与讨论
石鳖基因组显示出频繁且极端的重排
四个新的石鳖基因组代表了该纲中最完整的基因组,加上了之前发表的几个部分或完整的基因组,进一步增加该类群基因组的覆盖范围。我们测序了来自西太平洋日本大黑热泉(Daikoku vent field)的Deshayesiella sirenkoi(侧鳞石鳖目),来自北爱尔兰斯特朗福德湖潮间带的Callochiton septemvalvis(Callochitonida)和Acanthochitona discrepans(石鳖目),以及来自中国青岛潮间带的同属A. rubrolineata。这些材料首先进行PacBio HiFi测序,并通过Hi-C进行scaffolds组装,并得到了高质量的组装, BUSCO完整性超过97%(Callochiton septemvalvis为94%),各属的染色体数量在8-13之间不等(图1,附录,图S1,表S1)。这些物种表现出高水平的杂合性,从Deshayesiella的几乎1%,到Callochiton的4.12%不等(图1,附录,图S2)。
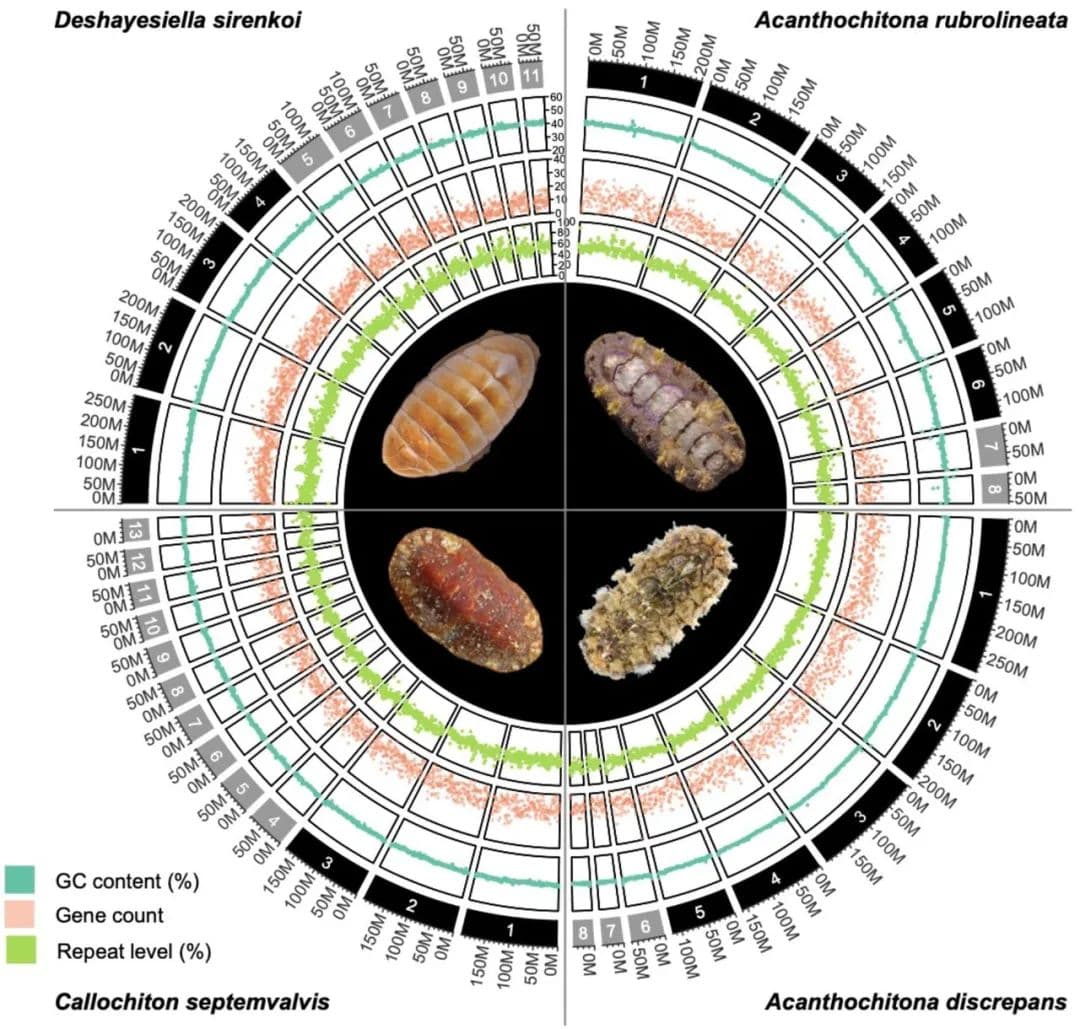
图1 四份新组装的石鳖基因组特征
我们的系统发育结果确认了石鳖作为单系有壳亚门的姐妹群的位置,并且我们恢复了与其他近期基于基因组和形态学特征研究一致的多板纲内的预期拓扑结构(图2)。在有壳亚门中,我们确认了其他近期研究的拓扑结构;然而,我们的补充分析发现了弱支持的掘足纲(Scaphopoda)与腹足纲(Gastropoda)姐妹关系(附录,图S3)。与其他软体动物类群的基因组比较显示,软体动物祖先的基因组由20个连锁群组成(图2),我们称之为软体动物连锁群(MLG)1-20。这确认了之前的研究,这些研究也基于其他后生动物预测了软体动物祖先的单倍体核型为20。三次重要的融合事件是多板纲明显的共有特征,存在于所有现存石鳖中,而不存在于有壳亚门中:MLG 4+16+18、MLG 7+10和MLG 8+9(图3,图4,附录,图S4-S7)。每个采样物种中还有其他的融合事件和染色体内重排。
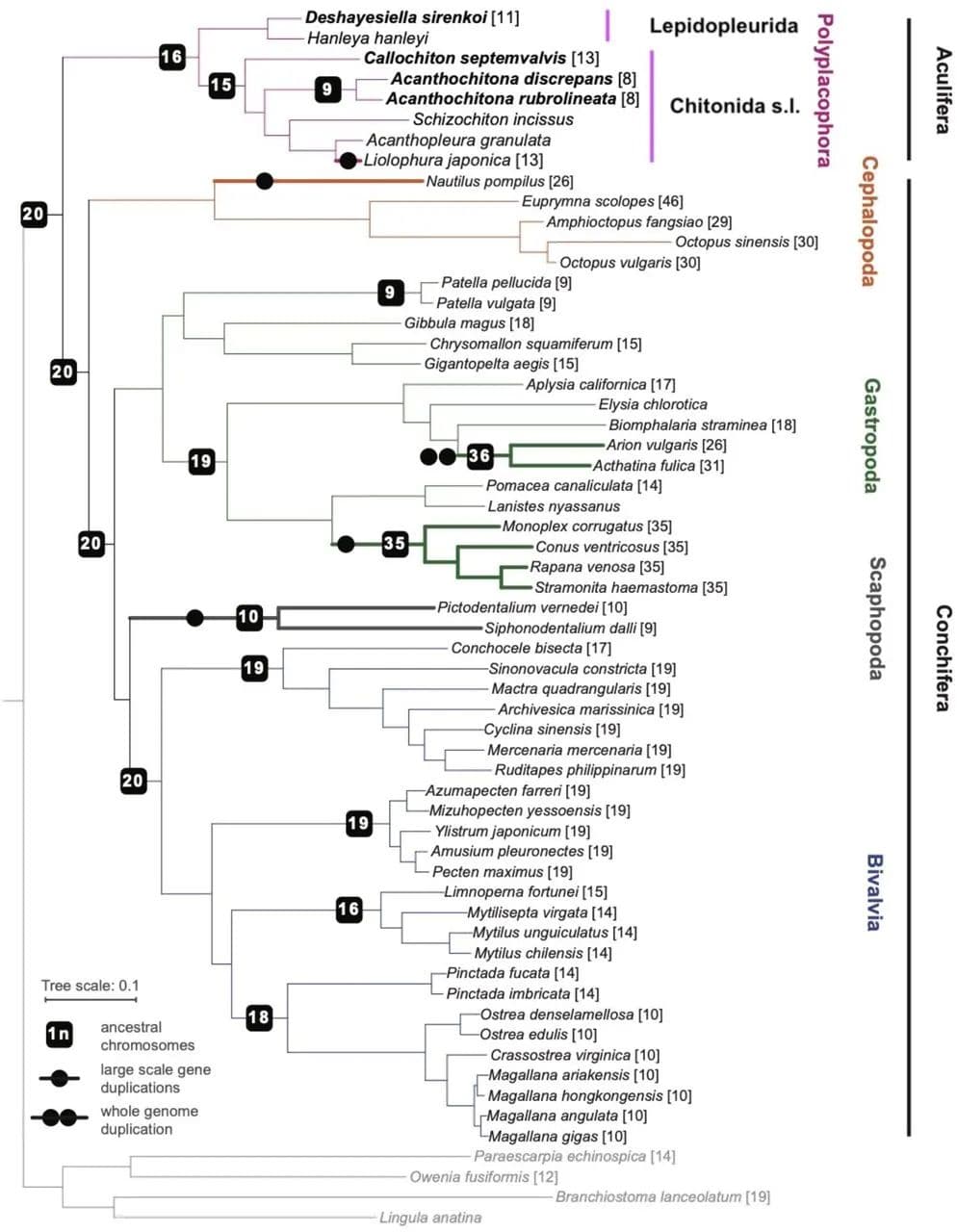
图2 软体动物系统发育关系
先前的研究展示了石鳖核型的可变性;石鳖目类群中的物种单倍体染色体数量从6到16不等,模式为11条。我们的数据首次指出,侧鳞石鳖目物种的预期染色体数量(1n=11)处于这个范围内。使用五个石鳖的染色体级基因组组装,我们重建了多板纲(更严格来说是分类学上的新石鳖目(Neoloricata))的祖先核型,并展示了石鳖进化过程中基因连锁群逐步融合和重排的各个中间系统发育节点(图3)。
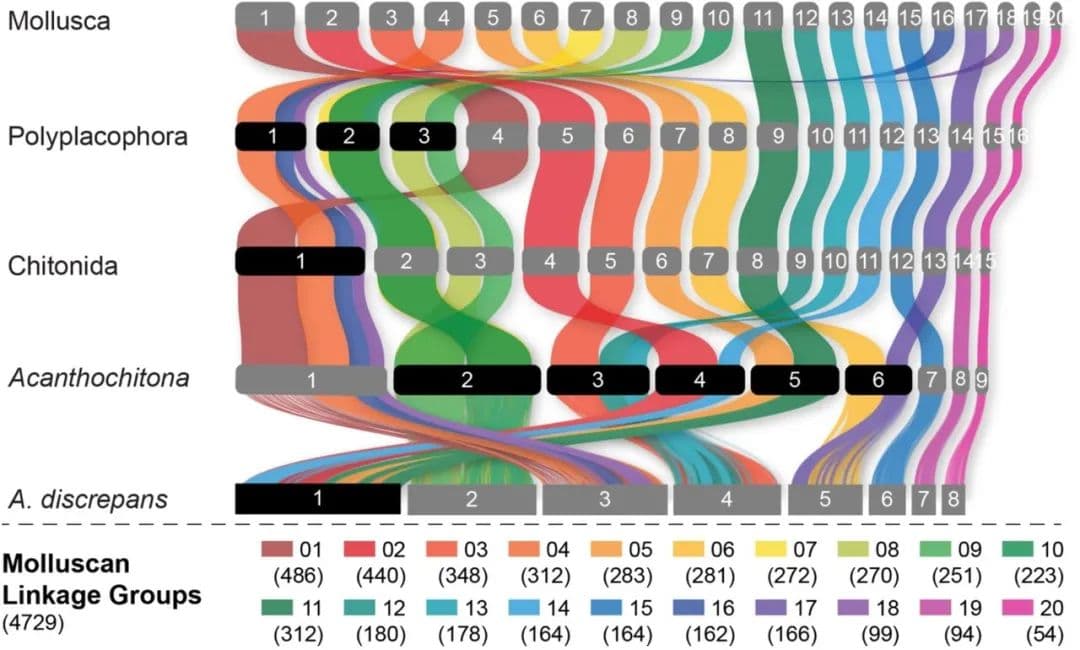
图3 石鳖与其他软体动物的比较基因组学分析(基于MLG)
即使在单一属内,石鳖展示了极端的基因组重排。这不仅代表着基因顺序的差异,还代表基因组结构的共线性变化。基于化石记录,Acanthochitona属物种的分歧时间相对较短,最大约为2300万年。Acanthochitona discrepans和A. rubrolineata各自有8条染色体(单倍体),但这些染色体来源于与重建的Acanthochitona祖先核型相比的两次不同融合(图1,图4C)。之前A. discrepans的核型数据实际上是基于A. crinita的标本;这两个物种非常相似,但可以根据地理分布进行正确鉴定。Acanthochitona crinita有9条染色体(单倍体):这是同属内的又一种共线性排列。在不同海洋盆地(太平洋的A. rubrolineata)之间,以及在大西洋东北处的两个物种(A. discrepans和A. crinita)之间存在重大变化,后者在形态和生态上几乎无法区分。
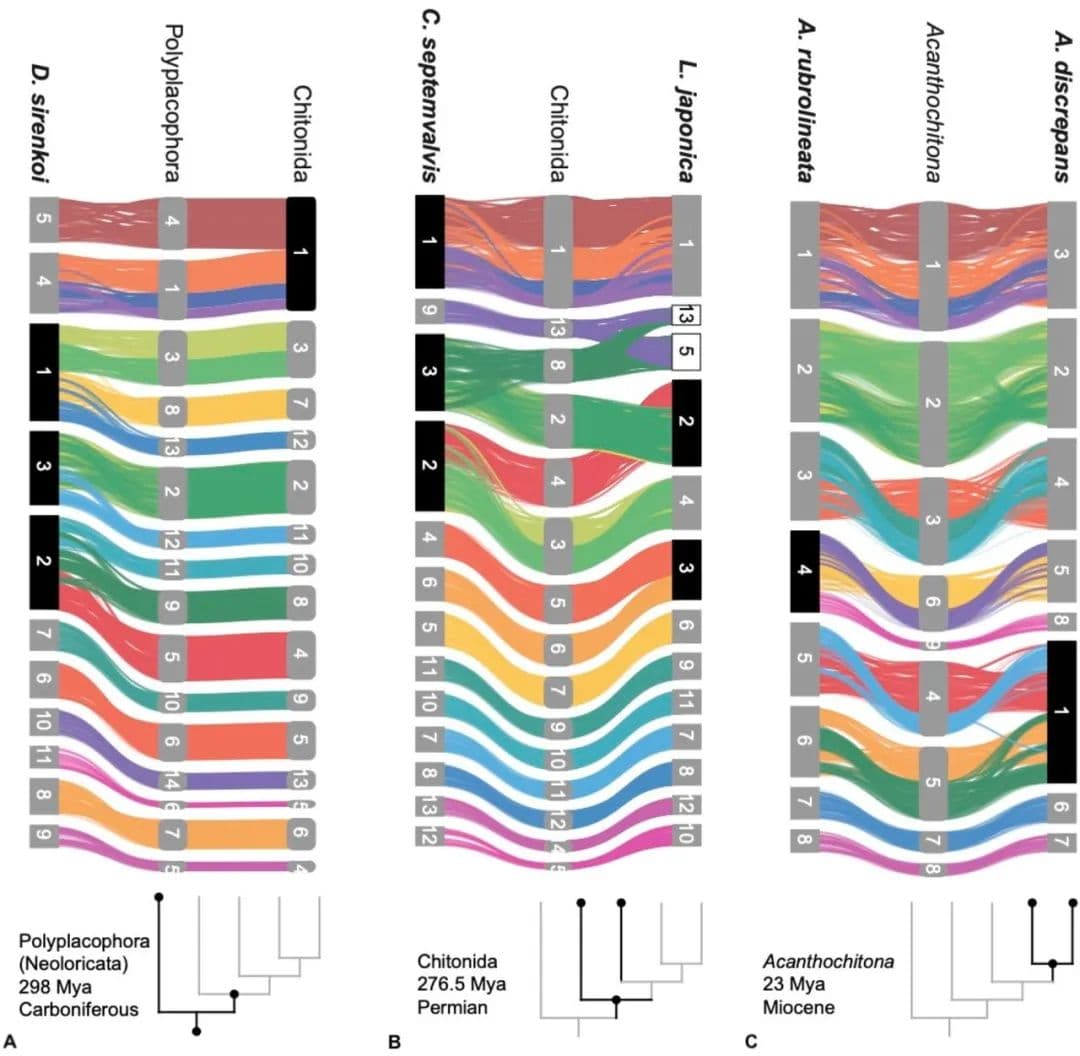
图4 在多板纲进化过程中,MLGs中共线性的重排
现存的侧鳞石鳖目物种保留了更多的原始形态特征,该类群的化石记录延伸到下石炭纪。在石鳖基因组中,侧鳞石鳖目Deshayesiella中有最大的三次新融合(图4A)。剩下的现存石鳖形成了两个姐妹类群,分别被认为是不同的目:石鳖目和Callochitonida。Callochiton也发生了两次额外的融合事件,而石鳖类Liolophura japonica有部分基因组重复,两个连锁群首先融合然后又重复(图4B)。在我们的祖先核型重建中,石鳖目的祖先与(石鳖目+Callochitonida)的祖先之间的排列没有差异。石鳖目(广义,包括Callochiton)的四个物种共享一个大的融合染色体(MLG 01+04+16+18),这在Liolophura japonica中明显没有很好地混合(即Liolophura japonica中的Chr01,Callochiton septemvalvis中的Chr01,Acanthochitona rubrolineata中的Chr01和A. discrepans中的Chr03)。这种模式的一部分起源于祖先石鳖核型,并在侧鳞石鳖目Deshayesiella中保留(即MLG 04+16+18)。这意味着染色体内重排的速率可变,其中几个MLG是保守的。
软体动物的祖先核型
动物的染色体数量并不高度保守。染色体数量的变化被认为是鳞翅目多样化的重要驱动因素之一,研究表明染色体数量变化速率与物种形成速率之间有很强的相关性。鳞翅目染色体数量的不稳定性主要通过核型数据进行研究,但最近通过基因组数据同样得到了确认,在属内的变化已知涵盖了中性进化和适应性进化。然而,这种重排的水平可能要比多板纲的要低。
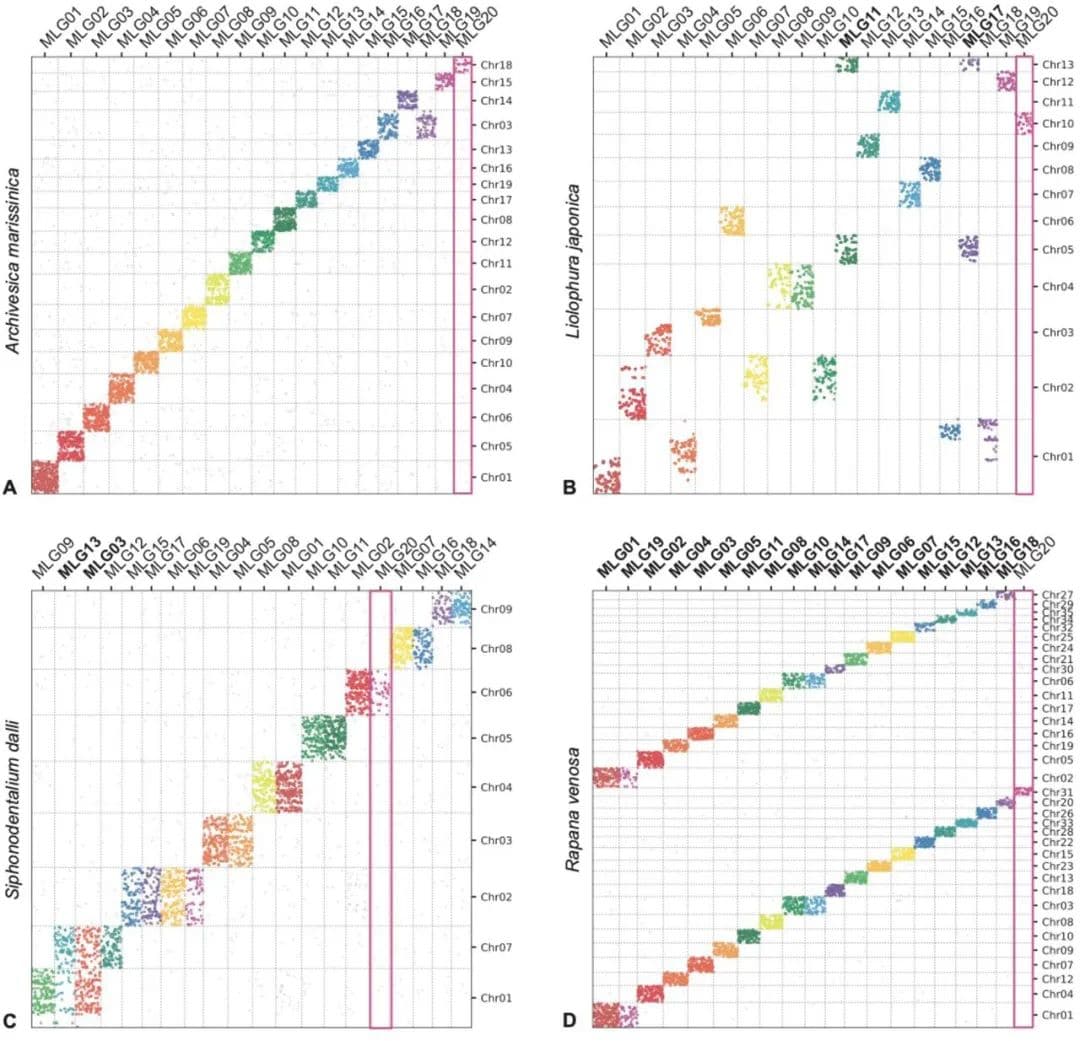
图5 基于MLGs的牛津点阵图
即使在单一物种内,染色体融合也并不罕见,罗伯逊易位在大约每1000个人中发生1次。先前的一项研究推测,近缘类群的石鳖染色体丢失可能是罗伯逊易位的结果。尽管我们仍然缺乏有关端粒的信息,但这一简单机制不足以解释几种更复杂的染色体内重排事件(如MLG 7+10: 图4)。染色体融合不会立即产生生殖障碍,但姐妹物种之间的染色体重排可以作为Dobzhansky-Muller不相容,并产生生殖隔离。石鳖是广播产卵者,因此这种障碍可能在是有利的,但没有发现这些共线性重排的双壳纲中也是如此。
为了比较多板纲内的共线性与其他软体动物类群的差异,我们重新分析了所有可用的软体动物基因组,以新识别的MLG为背景。例如,双壳纲Imparadentia(Lucinida + Venerida)类群的分歧时间估计在志留纪,大约4.3亿年前,比冠群多板纲的分歧时间早了超1.5亿年,但Imparadentia内的物种几乎没有共线性重排(见附录,图S7,S8)。起源于更早的奥陶纪下层(大约4.85亿年前)的双壳纲Pteriomorphia显示出一些重排,但两个目(Arcida和Pectinida)的成员高度相似,而两个被广泛养殖的牡蛎目(Ostreida)物种与其他物种以及彼此之间存在差异(见附录,图S8)。
潜在的适应性作用可能与不同MLG的相对移动性有关。五个石鳖基因组中最小的两个MLG是最保守的;MLG20是唯一在所有双壳纲和石鳖类群中保持独立的。该区域在Neogastropoda的部分基因组重复中也没有重复,但在两种雌雄同体陆生腹足类(Achatina fulica和Arion vulgaris)的全基因组重复中被复制(图5,见附录,图S5)。
基于核型数据的早期模型预测,在现存软体动物的进化历史中有三次全基因组重复事件:在Neogastropoda、Stylommatophora和有壳头足类。Stylommatophora类群的可用基因组证实了全基因组重复的存在,以及Neogastropoda类群中的大规模基因重复(见附录,图S6)。在有壳头足类基因组中,MLG和密集融合的共现可能不容易解决,但更有可能源于先前工作中报告的染色体碎裂过程。相比之下,在鹦鹉螺中(Nautilus),包含保守Hox基因集的连锁群(Chr11, MLG16),没有显示出与其他MLG或其他染色体共现的信号(见附录,图S6)。新的结果还显示了石鳖Liolophura中的部分和古老的基因组重复(图2,图4)。鉴于MLG在所有石鳖中都具有良好保守性(如果不是它们的顺序)(见附录,图S4),Liolophura japonica中的事件很可能是重复而不是裂变。现已知有四个软体动物纲中的全基因组或部分基因组重复,表现出不同的倍性模式(如腹足类)或串联重复(如掘足类)(图5)。
重建基因组进化对于具有高水平染色体内重排的类群自然更为困难。相比于基因顺序变化的更快速率,这些在相同染色体上共现的基因位点(共线性)的模式,应该在进化时间上保持更长时间。共线性变化不一定遵循总体易位率,不同软体动物类群之间的差异并不明显(见附录,表S4,S5)。快速基因顺序或连锁群变化的混淆效应不仅限于系统发育距离的问题,即使是近缘的物种也存在显著的共线性差异时也是如此。尽管如此,关注连锁群排列是解决许多深层软体动物系统发育未解问题的一个富有前景的方向。最近的研究强调了基于共线性研究基因组进化的重要性,这是作为理解深层分歧以及基因组结构、融合、裂变和易位基本机制的基础。昆虫和脊椎动物的一般趋势是衍生谱系中染色体数量的增加。在最近的石鳖研究中,融合事件可能更为常见,这可能与特定的适应有关。
理解多样化需要多样化的类群
石鳖是广播产卵者,不迁徙,不太可能受到强烈的性选择影响,许多相似物种在近邻或同域辐射中共存(parapatric or sympatric radiations)。染色体重排是一个具有吸引力的推测性解释,可能起到维持这一类群中物种界限的作用,例如石鳖目Acanthochitona discrepans(1n=8)和A. crinita(1n=9)。对于大多数软体动物类群来说,参考质量基因组数量相对较少是限制这些分析的重要因素。高杂合性在软体动物中似乎非常常见,显然是石鳖的一个特征,这给高质量基因组组装带来了困难。Callochiton的杂合率为4.12%,远远超过报道的“最高之一”的鳞翅目基因组杂合率2.95%。基于基因组学的大规模分析对遗传多样性的驱动因素有不同看法,因此需要包括来自不同类群的更多数据。
一个关键问题是多板纲软体动物本身是否保守,或者其特有的躯体构造适应性(diagnostic body plan adaptations)如此独特和奇怪,是否以至于掩盖了对其他适应性特征的全面理解:完全受神经支配的壳(fully innervated shells)、铁矿化齿舌(iron mineralised radula)、并且生活在几乎所有深度和纬度的海洋中。尽管石鳖的身体构造已持续保持超3亿年,但这是一种模板,体现了近年来才开始被重视的非凡的适应性。
“多样性群体”(“diverse groups”)这一概念主要基于感知到的变异性,通常表现为形态、生态或遗传差异。鳞翅目通常被认为是超级多样化的类群,但它们引人注目的颜色和不同的翅膀形状是一个明显受限主题内的变异。石鳖尽管存于不同生态位中,物种数超过1000种,具有广泛的形态适应性,过去却被认为是一个“次要”和“被忽视”的类群。该类群在分类学上具有挑战性——一方面,由于作为一个类群被假定认为处于形态停滞,另一方面,由于其高的种间变异性。石鳖表现出了染色体重排,甚至在Acanthochitona属内,重排达到了几乎前所未有的水平。这在首次比较基因组研究中得到了明确体现,相对于其他研究的类群有更多的数据积累。所有这些都提出了一个更普遍的问题,即我们如何定义变异性,我们如何感知它,以及我们是否真正理解它?尽管鳞翅目的变异性显而易见,石鳖同样是多变的,这可以通过其基因重排得到清晰展示。在被忽视的生物类群中,认识到基因组和表型的变异性对于不偏倚地理解和重新评估多样性的衡量标准至关重要。
通讯作者

孙进,中国海洋大学海洋生物多样性与进化研究所教授、博导,国家优秀青年基金(海外)、山东省泰山学者青年专家获得者。
先后在中国海洋大学、香港浸会大学获得学士、博士学位,后加入香港科技大学钱培元教授课题组。
学习工作简历
2020年11月加入中国海洋大学。主要研究方向是深海生物的适应性进化,累计在《Nature Ecology & Evolution》、《Nature Communications》等领域顶级刊物上发表论文80多篇。任《The Innovation》杂志青年编委,《Frontiers in Marine Science》杂志副编辑。
曾以首席科学家或科考队员身份多次参加日本JAMSTEC冲绳热液区调查航次、中国大洋航次西南印度洋热液区调查航段、中国南海冷泉调查航次等。
主要研究方向简介
主要研究领域为以分子和组学为基础的进化生物学及环境生物学,聚焦深海热液、冷泉等化能合成环境中动物与微生物的共生关系、适应性以及软体动物物种多样性的维持机制。主要成果包括:深海极端生命的适应性进化机制及群体连通性;生物矿化机制及进化历程;入侵生物-福寿螺的物种演化及进化途径。
迄今上述成果累计在(包括《Nature Ecology & Evolution》、《Nature Chemical Biology》、《Nature Communications》(4篇)、《Molecular Biology & Evolution》、《ISME Journal》、《Microbiome》等领域顶级)刊物上发表论文80多篇。
CITATION
Still waters run deep: Large scale genome rearrangements in the evolution of morphologically conservative Polyplacophora
Julia D. Sigwart, Yunlong Li, Zeyuan Chen, Katarzyna Vončina, Jin Sun
bioRxiv 2024.06.13.598811; doi: https://doi.org/10.1101/2024.06.13.598811
信息来源 | 进化随想微信公众号
转载自智慧海洋圈子




